
Нещодавнє зростання досліджень епігенетичних механізмів, описаних у низці публікацій 2014 року (Noble et al., 2014), дало змогу розширити поняття неодарвіністського синтезу.
На наш погляд, потрібно говорити про заміну категориального апарату сучасної теорії еволюції. Причина в тому, що існування надійних механізмів трансгенного успадкування, незалежних від послідовностей ДНК, повністю суперечить духу модернового синтезу. Деякі нові особливості експериментальних даних щодо успадкування та механізмів еволюційної мінливості несумісні з базовими теоретичними положеннями.
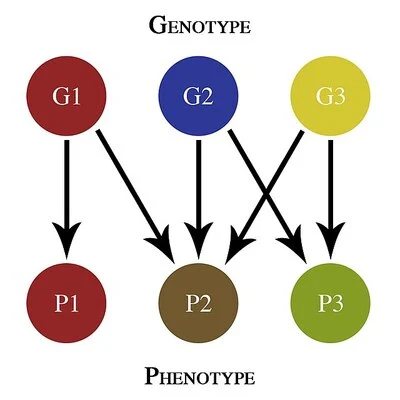
 На малюнку продемонстровано дефініції та взаємозв'язки між різними ознаками дарвінізму, модернового синтезу і пропонованого нового інтегративного синтезу. Діаграма ґрунтується на логіці, запропонованій Пінліуччі та Міллером у 2010 році, що пояснює ідею розширеного сучасного синтезу.
На малюнку продемонстровано дефініції та взаємозв'язки між різними ознаками дарвінізму, модернового синтезу і пропонованого нового інтегративного синтезу. Діаграма ґрунтується на логіці, запропонованій Пінліуччі та Міллером у 2010 році, що пояснює ідею розширеного сучасного синтезу.
Перехід до нового синтезу в еволюційній біології також можна розглядати як частину більш загального зсуву точки зору в бік більш системних підходів. Редукціоністський підхід (який розуміється як генно-центрована теорія еволюції) був дуже продуктивним, але його завжди потрібно доповнювати інтегративним підходом, зокрема новою теорією причинності в біології.
Деякі дослідники навіть заговорили про виникнення теорії біологічної відносності за аналогією з ейнштейнівською концепцією. Сучасний підхід можна назвати інтегративним синтезом, оскільки він ґрунтується на інтеграції різних механізмів еволюційних змін, а не на вірі, постульованій неодарвіністською логікою. Тобто йдеться про N-елементну теорію еволюції, яка, що цікаво, ближча до деяких ідей Дарвіна, ніж до неодарвінізму. Зокрема, йдеться про визнання інших, крім природного добору, механізмів еволюції, - наприклад, успадкування набутих ознак.
Мова неодарвінізму
Багато з проблем сучасного синтезу, які полягають в акомодації нових експериментальних даних, походять із неодарвіністського уявлення, а не з самої експериментальної бази.
Ці форми репрезентації виражають інституційні особливості біології 20-го століття в академічній науці. Отже, ми маємо розглянути методи інтерпретації фіксованих дослідів. А також зважати на ту обставину, що мова неодарвінізму відображає вкрай редукціоністські погляди, які не потребують наукових відкриттів як таких.
 Особливо це помітно на прикладі операціоналізації таких понять, як "егоїстичні гени" або "генетична програма". Концепції набувають упередженої інтерпретаційної оболонки, яка здатна приховувати відкриття в мережі нагромаджених інтерпретацій.
Особливо це помітно на прикладі операціоналізації таких понять, як "егоїстичні гени" або "генетична програма". Концепції набувають упередженої інтерпретаційної оболонки, яка здатна приховувати відкриття в мережі нагромаджених інтерпретацій.
Непевні тлумачення в неодарвінізмі продукують потік спекуляцій, відірваних від експериментальної науки. Точніше навіть так: не кожна теорія потребує підтвердження, але кожен досвід породжує кілька істин, що існують одночасно, в імовірнісному порядку, зрозуміло.
Оскільки сучасний синтез домінував у біологічній науці впродовж більш ніж півстоліття, його методологічні позиції настільки вбудовані в наукову літературу, включно зі стандартними шкільними та університетськими підручниками, що багато вчених-біологів можуть не усвідомлювати концептуальної природи науки, якою вони займаються. Сумніватися в теорії для вченого означає піддатися гріхопадінню. Часто академічні протести закінчуються вигнанням із наукового середовища.
Тому на виході ми бачимо лише опис експериментів; ви рідко дочекаєтесь аналізу, узагальнення, а тим паче теоретизування проблеми.
Приміром: гени кодують білки, які формують організми за допомогою генетичної програми, успадкованої від попередніх поколінь, і які визначають, як будуть "організовані" їхні нащадки. Що не так із цим положенням?
Ген
Неодарвінізм - це генно-центрована теорія еволюції. Проте, його центральне поняття, "ген", є нестабільною категорією. Як не дивно, єдиного узгодженого визначення цього поняття не існує. Ба більше, різні визначення мають несумісні наслідки для теорії.
Слово "ген" було введене Вільгельмом Йоганнсеном для визначення спадкового фактора. Але сама концепція вже існувала за часів Менделя і стосувалася дослідів на гібридних рослинах.

Але якщо всі фенотип-характеристики не можуть бути співвіднесені виключно до ДНК-послідовностей (ДНК не діє поза контекстом повної клітини), то ген М не є таким самим, як ген J.
"Егоїст"
Не існує біологічного експерименту, який міг би провести відмінність між егоїстичною теорією генів та її протилежностями, такими як "ув'язнені у в'язницю" або "кооперативні гени". Цю точку зору давно визнав Річард Докінз у своїй книжці "Розширений фенотип": "Я сумніваюся, що існує будь-який експеримент, який міг би довести моє твердження".
Код
Після відкриття подвійної спіральної структури ДНК було виявлено, що кожна послідовність із трьох основ у ДНК або РНК відповідає одній амінокислоті в послідовності білка. Ці триплетні патерни формуються з будь-якої комбінації чотирьох основ U, C, A і G в РНК і T, C, A і G в ДНК. Їх часто називають генетичним "кодом", але важливо розуміти, що таке тлумачення несе в собі обертони, які можуть збивати з пантелику.
 Код спочатку був навмисним шифруванням, використовуваним людьми для спілкування. Генетичний "код" не є навмисним у цьому сенсі.
Код спочатку був навмисним шифруванням, використовуваним людьми для спілкування. Генетичний "код" не є навмисним у цьому сенсі.
Річ у тім, що самі клітини визначають патерни експресії для різних станів. Як сказала лауреатка Нобелівської премії Барбара МакКлінток, геном - це "орган клітини", а не навпаки.
"Програма"
Ідея "генетичної програми" була запропонована французькими нобелівськими лауреатами Жаком Моно і Франсуа Жакобом.
 Якщо так станеться, то через десятиліття ми навчимося, як лікувати рак, хвороби серця, нервові захворювання, діабет.
Якщо так станеться, то через десятиліття ми навчимося, як лікувати рак, хвороби серця, нервові захворювання, діабет.
Література
Beurton, P. J., Falk, R. and Rheinberger, H.-J. (2008). The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives. Cambridge University Press.
Джерело: Evolution beyond neo-Darwinism: a new conceptual framework










